Maintenance of variation within populations
We are also using Penstemon species to study the genetic and ecological mechanisms for floral variation within (and across) populations.

|
We seek to understand how genetic and ecological processes generate and maintain trait diversity. We are primarily exploring these issues in Penstemon, an enigmatic comparative system for studying complex trait evolution. Our work developing Penstemon as a model for evolutionary genetics is collaborative with Lena Hileman and John Kelly.

|
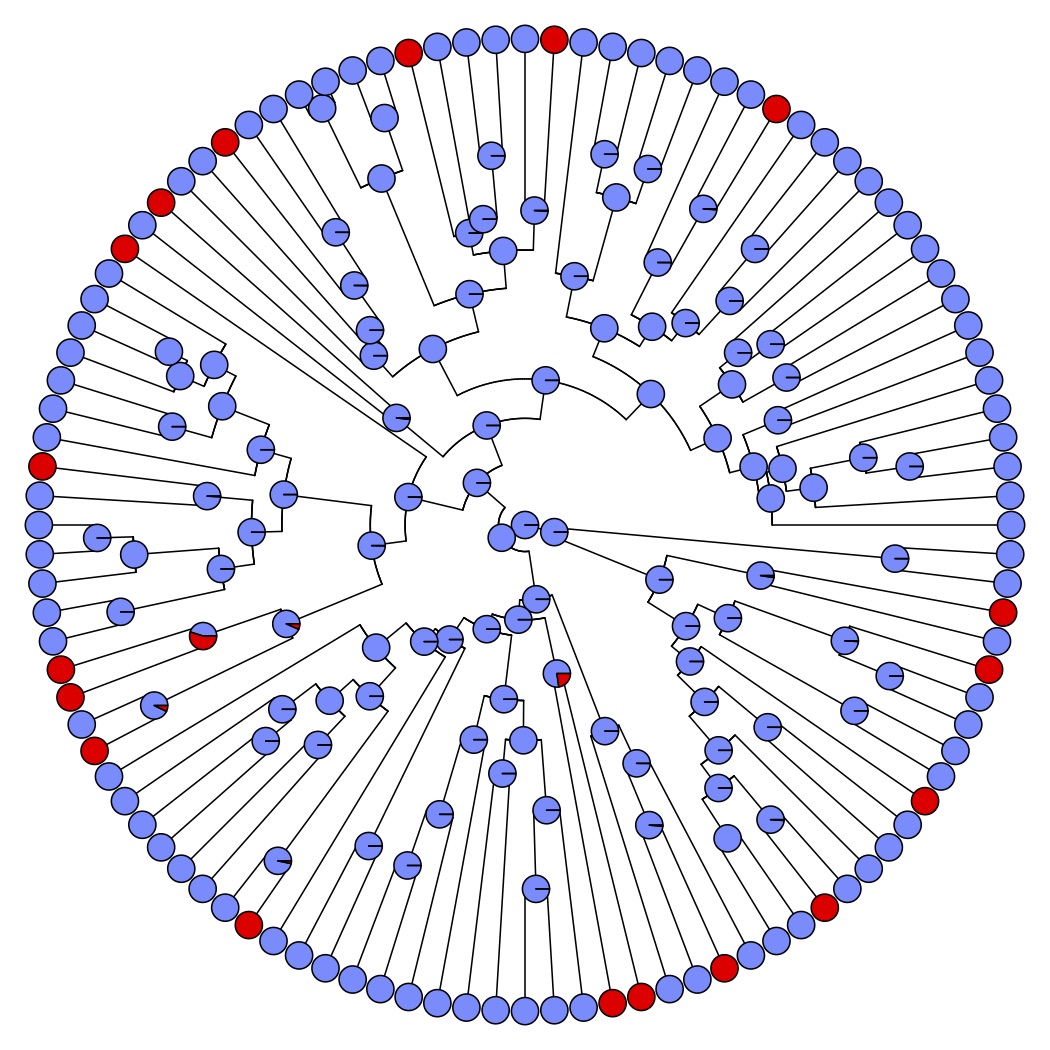
In the North American wildflower genus Penstemon, evolutionary transitions from bee to hummingbird pollination have occurred in an estimated 20+ lineages through changes to a complex of floral traits. Recent macroevolutionary analyses suggest that transitions to hummingbird pollination are associated with reduced speciation rates and that reversals back to bee pollination are rare.
We view Penstemon as an exceptional comparative system to study fundamental evolutionary genetics questions, using approaches such as population genomics, quantitative genetics, comparative studies, phylogenomics, and modeling.
|
Hummingbird-adapted flowers have evolved a surprising number of times in separate bee-adapted lineages! This suggest the “hummingbird syndrome” strategy is often favored by selection and is genetically accessible to populations. Does standing variation or gene flow act as a source of adaptive alleles? In theory, adaptive introgression could bypass the waiting times for relevant de novo mutations. We are using whole genome resequencing and phylogenomic approaches to address whether hummingbird-adapted species share a history of introgression in genome regions that specify floral traits.
A critical goal of evolutionary genetics is to characterize the types of mutations that contribute to adaptive trait variation. We are using QTL mapping to examine the genetic basis of repeated shifts to hummingbird pollination using replicate mapping populations. What components of parallel phenotypic evolution occur through similar genetic changes?
In addition, we are studying the ecological function and genetic basis of personate flowers within Penstemon, using field studies and QTL mapping. Personate flowers are dorso-ventrally flattened, impeding pollinator visitation relative to open tubular flowers. Personate flowers have arisen at least twice within Penstemon, suggesting they have adaptive significance. However, little is known about how and why this interesting flower shape has evolved!

|
Evolutionary change in complex traits begins with local adaptation within populations. A major challenge is understanding how complex trait differences are maintained in recently diverged taxa that experience gene flow. We are using population genomic approaches to examine the heritability of complex adaptations within populations and to determine the consequences of gene flow on species differentiation. We are focused on recently diverged pairs of sister species that differ in floral syndrome and show evidence of gene flow where species ranges overlap.

|
|
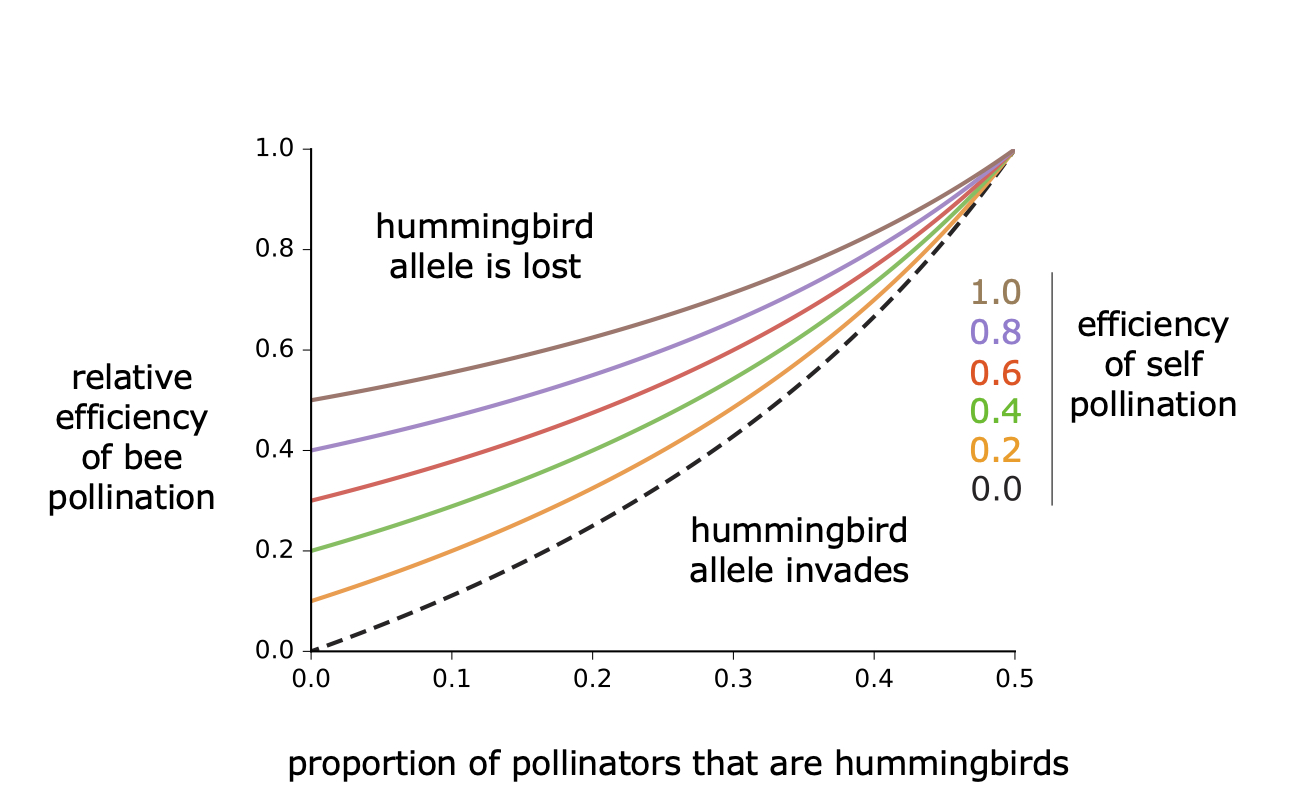
Mating system has fundamental effects on the genetics of populations, including the fate of new beneficial alleles. Adaptation to a new pollinator may alter patterns of pollen dispersal within and among populations, with effects on population isolation, local adaptation, and the occurrence of hybridization. We are studying the consequences of pollination system on mating events and population structure, which may have relevance for macroevolutionary dynamics.
We are also using Penstemon species to study the genetic and ecological mechanisms for floral variation within (and across) populations.
|
|